
What is the American Science and Security Cloud? Data Platforms Program Lead Dylan McReynolds explains how the ALS is using this network of tools and helping build an even more robust platform. Read more »
Powering Science: How SYNAPS-I Builds on Meta’s Open Source AI
SYNAPS-I, a DOE initiative led by Berkeley Lab, unites 60 researchers across five labs to speed up X-ray and neutron data analysis using AI. Because prepublication data must stay on secure government systems, open-source models from Meta — SAM 3 and DINOv3 — let the team fine-tune cutting-edge AI entirely in-house. Read more »

The ALS-U Project Progresses with New Director
The Advanced Light Source Upgrade (ALS-U) project has received approval for a revised cost and schedule, targeting completion by late 2029. Greg Hays joins the team to steer the historic modernization project, which will increase soft X-ray brightness a hundredfold and enable AI-driven research in materials science, quantum computing, biology, and more. Read more »

ALS-United: Tianhuan Luo and David Nett
ALS-United is an opportunity to meet the people collaborating at the Advanced Light Source and the ALS Upgrade Project. Hear firsthand how team science enables the cutting-edge research of today and builds the facility of the future. This month, we spoke with Tianhuan Luo (Research Scientist) and David Nett (Electronics Engineer). Read more »
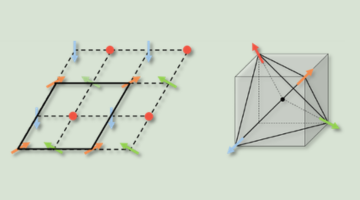
Revealing the Electronic Signature of Unusual Magnetism
Experiments at the ALS showed how magnetic Co atoms sandwiched between TaS2 layers reshape the material’s electronic structure. Understanding how unusual magnetic order influences electron movements in new quantum material CoxTaS2 could guide its use in advanced quantum technologies based on spintronics and valleytronics. Read more »